




服务热线
400-998-2324

前体蛋白是无活性的,需要通过翻译后修饰(PTM)才能解锁繁多的蛋白类型和功能。翻译后修饰(PTM)是调节蛋白质功能的一种极其重要的机制.目前,已知在哺乳动物细胞蛋白中发现了600多种修饰。除了磷酸化、糖基化、乙酰化、泛素化和甲基化等经典的PTMs外,越来越多的新的修饰被发现,如琥珀酰化、羟丁基化和乳酸酰化等,在人类癌症的治疗和预防方面的应用前景有待进一步探索。
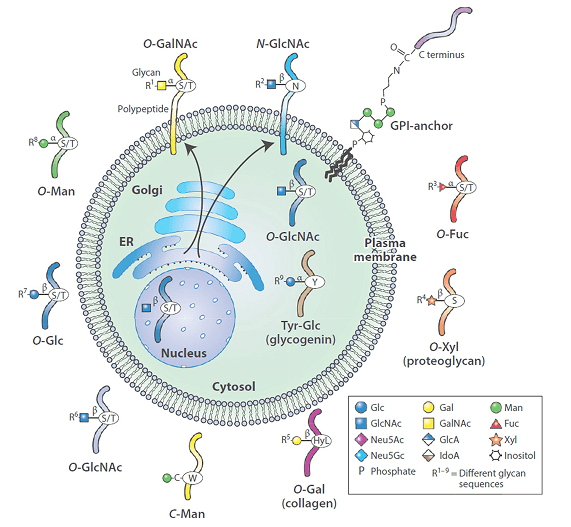
哺乳动物的糖基化主要有两种类型,以天冬氨酸连接(N-连接)或丝氨酸/苏氨酸连接(O-连接)寡糖形式存在的碳水化合物是许多细胞表面和分泌蛋白的主要结构成分。蛋白质糖基化的改变可以改变蛋白质的构象、寡聚和转换,从而导致细胞信号通路的改变。
下图展示的是细胞膜和分泌蛋白的糖基化
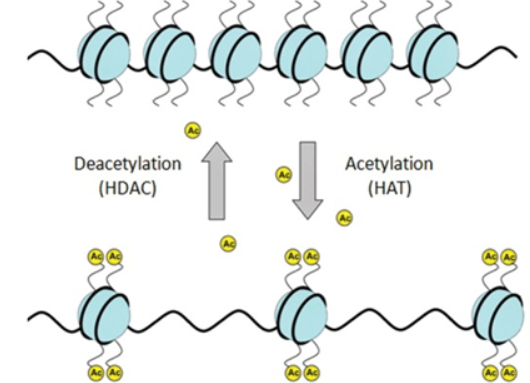
乙酰化修饰分为2类:蛋白N端的乙酰化修饰和蛋白赖氨酸的乙酰化修饰。80- 90% 的真核细胞蛋白是N-乙酰化,但其确切的生物学意义仍不清楚。
早期,乙酰化研究主要集中在细胞核内的组蛋白以及其对基因转录表达调控上。许多转录辅助活化因子都具有乙酰基转移酶内在活性,而转录辅助抑制因子则与去乙酰酶活性相关。后来,随着质谱技术的发展,发现在胞质或其他细胞器中大量的非组蛋白存在赖氨酸乙酰化修饰的现象。越来越多的非组蛋白的特定位点乙酰化已被证实可调节其活性、定位、特异性相互作用以及稳定性/降解。
乙酰化是一个普遍而重要的蛋白质翻译后修饰,不仅集中在对细胞染色体结构的影响以及对核内转录调控因子的激活方面,而且还影响参与细胞周期和新陈代谢、肌动蛋白聚合控制、肿瘤及多聚谷氨酰胺疾病等方方面面。蛋白质乙酰化也是近年来众多疾病新药开发设计的有利靶标。

品质保证
10年以上科研试剂研发经验
.

专业实力
抗体试剂近上万种
专业试剂研发团队

售后保障
厂家现货直发
24小时技术在线

定制服务
提供科研试剂所需数据,按需定制
订货周期短,确保实验准确性